2.1 両眼間の対応
両眼間の対応感度とステレオ視の関係
弱視者は両眼視融合をもつがステレオ視ができないことが知られている。健常者でも一定の視差条件での左右眼の対応度はステレオ視の強さの測度となる(Cisarik & Harwerth,2008; Doi, et al., 2011; Henriksen, et al.2016a; Henriksen,et al. 2016b; Julesz & Tyler, 1976; Tyler & Julesz, 1980)。Reynaud & Hess(26)は、ステレオ視と両眼間の対応関係を両眼間の刺激対応度を操作して検討した。図5には、刺激の両眼相関関係(IOC、interocular corrlation)と視差対応の各パターンが示されている。図(a)には両眼間の刺激相関の操作に用いたパターンを示した。左右眼に提示するパターン(llとlr)は位相を異にするサイン波形の包絡線(ma,mc、1/4周波数)をもつノイズパターンで変調した2種類のノイズテクスチャ(ccとcl、 ccとcr)をブレンドして作成され、ccは両眼間で共通のテクスチャをもち、また両眼間のイメージ(llとlr)に共通するストライプパターンの相関の程度は変調パラメータ(m)によって規定された。図(b)には両眼間の視差パターンを示し、それらは同様なノイズ刺激から構成され、水平視差の変調はノイズ刺激の1/4周波数をもつ斜方向の波形で規定された。IOCの感度は融合視させたパターンの刺激の方向を、また視差感度も立体視させた刺激パターンの方向(45°または135°)を、それぞれ被験者に答えさせる方法て得た。この場合、刺激パターンの空間周波数は0.24cpdから2.39cpdの範囲で8段階に、また両眼間のIOCは0.10から1.00の範囲で11段階にそれぞれ変えられた。さらに刺激持続時間(0.05、0.1、0.15、0.25、0.5、1、2、3 秒)および両眼間の刺激コントラスト(0.3、0.5、0.7、1.0)も操作された。実験では刺激はLEDモニターに提示し、6名の健常眼被験者には偏光グラスを装着させ両眼視させた。
実験の結果、左右眼別々にサイン波形をもち左右眼での刺激対応度を変調させた帯域通過型のテクスチャ刺激に対するIOC感度は、空間周波数の低帯域通過型あるいは帯域通過型の関数特性を示した。また左右眼の対応度を変調させた視差感度も同様な関数特性を示した。これらのIOC感度と視差感度は刺激持続時間に伴って向上し、とくに高空間周波数パターンで顕著に示された。刺激コントラストを操作した場合の空間周波数に伴うIOC感度は刺激パターンのコントラストが0.3のときにもっとも悪く0.5-1.0の条件で増大した。空間周波数に伴う視差感度はコントラスト1.0でもっとも良く、低下するにつれ悪くなった。さらに、IOC感度(測定値の計算によって求められた最大感度)と視差感度(測定値の計算によって求められた最大感度)の各被験者データ間の相関を求めると相関係数は0.137、また空間周波数でみた最大IOC感度と視差感度の相関は0.207となった。これらはいずれも有意な正の相関を示した。しかし、左右の眼球バランスの各被験者の測定値(0.7から1の範囲)と各被験者のIOC 感度あるいは視差感度をしらべると、眼球間バランスと2つの感度の間には優位な相関は示されなかった。
これらの結果から、IOC 感度は弱いながらも視差感度と相互に関係ていた。このことから、両眼視機能の障害をもつ弱視者では、IOC感度は視差感度よりも精度の高い測定方法として使えるかもしれない。
ステレオ刺激からの面のアモーダル(amodal)な統合に関わる知覚メカニズム
視覚システムは、部分的にオクルードされた3次元の面をいくつかの面の断片から統合して一つの面として知覚できる。部分から全体を知るこのようなアモーダルな面の統合は、オクルードする対象によって隠された各面の断片を外挿あるいは隠れた部分を内挿して全体の面としてまとめ上げることで成し遂げられる。図6には面のアモーダルな知覚統合が示されている。いま、2本の黒色縦帯刺激に隠された横長の白色長方形(a)を観察すると、その網膜イメージで白色面は3つの分割面に分けられる(b)。しかし中心の断面は背景より明るく(positive contrast-polarity)、左右端は背景より暗く(opposite contrast-polarity)すると、白色面は3つの断面に分けられる。 Su et al. (2010)によると、黒色の四角形刺激(静止)でオクルードされた同一のコントラストをもつ交差帯状パターンを矢印の方向に動かす(c)と、観察者は四角形内の帯状刺激が相互にスライドするように交差するのを知覚する(e)。これは四角形で隠された分割刺激を一つの運動刺激としてアモーダルに統合して知覚するためである。一方、交差する帯状パターンのコントラストが黒色四角形の内と外でコントラスト極性を反対(f)にとると、黒色枠内の交差帯状刺激は静止し、枠外の帯状刺激が拡大あるいは縮小して視える(g) 。これらは、オクルードされた2つの刺激が背景に対して一つのものとして捉えるか、あるいは3つのものとして捉えるかによって、それらを運動させたときのアモーダルな統合が異なり、その結果、観察者の知覚様相が相違するためである。これまでの多くの研究によると、3次元対象面の刺激特性である色、形状、空間関係、幾何的投影などの観察者の内的経験知識のいずれかに基づいてオクルードされた分割面がアモーダルにひとつに統合される(Su et al.2010a,b)。
He et al.(12)は、両眼立体視事態で視差をもつ面と視差ゼロの面が同一のあるいは異なるコントラストをもつ場合のアモーダルな統合の過程をしらべた。図7は実験に用いたステレオグラムで、(a)には、対象面である白色横帯を同一の輝度コントラストをもたせたステレオグラムを示し、図の左側2つステレオグラムは交差視差、右側2つは平行視差でこれらを両眼立体視すると白色横帯刺激を背後にもつ2本の縦帯刺激が出現する。図(b)には、中央が白色、左右端が黒色という反対コントラストをもつ横帯刺激と2本の縦帯刺激で、両眼立体視すると黒色横帯刺激を背後に左右端が灰色、中央が白色の縦帯刺激が出現する。これらのステレオグラムではいずれも白色横帯刺激の3つの領域のうち中央矩形の視差はゼロに、それを挟んだ左右端の矩形には5段階の非交差視差(0、2.9、5.7、8.6、11.5′)を付し、また中央矩形と左右端の矩形間のコントラストを白色あるいは黒色の2段階に設定した。観察はハプロスコープを用い、注視点のそばに提示した灰色ドットの視差を変化させ中央の矩形領域の奥行位置にマッチングさせて測定した。被験者は5名の成人であった。
実験1の結果、3つの矩形領域が同一のコントラストをもつ場合には両端の非交差視差の増大に伴って視差ゼロの中央矩形の視えの位置はリニアに増大したが、コントラストが異なる場合には中央矩形の視えの奥行位置はほぼゼロを示した。3つの矩形領域のコントラストが同一の場合には中央の矩形は左右端と同等の奥行に知覚され、一つの面としてアモーダルな知覚統合が起きていることを示した。しかし3つの矩形領域のコントラストが反対の場合には中央の矩形と左右端のそれらの奥行位置は異なり、アモーダルな統合は起きないことを示した。
一般に視覚システムは、相対視差による2つの面の出現に際し、それらの面の辺縁にある境界がどちらの面に所属するかを決める(Nakayama & Shimojo 1990a,b, Zhou et al. 2000)。図8に示したように、単眼視手がかりのT接合(T-junction)の境界は非交差視差を付した左右端の矩形ではなく縦帯刺激に属する(図のa)。一方、横帯の灰色断片は境界をもたないために、横帯の左と右の分割面は容易に広がってひとつになる(図のb)。その結果、横に広がった中央の矩形はコントラストも同一な左右端の分割面とつながりひとつの面を形成する(図のc) ので横帯の分割面はひとつに統合されて知覚される。
それでは、アモーダル統合過程において両眼視差のみがあれば2つの単眼視手がかりである境界効果がなくても複数の面が統合されるのであろうか。実験2では、実験1と様に、横帯刺激の中央矩形の奥行位置が測定された。図9はそのステレオグラムでコントラストが同一条件(a)と反対条件(b)でいずれも横帯の左右端の境界所属性が曖昧にしてある。図 (a)には縦帯の長さと横帯の幅を等しくし、コントラストは横帯の3つの矩形で等価にしたステレオグラム(立体視の出現様相はステレオグラムの下部)を示し、図(b)には両端の矩形と中央のそれとのコントラストを反対にしたステレオグラム(立体視の出現様相は下部)を示した。実験の結果、コントラストが同一条件では両端矩形の視差を大きくすると視差ゼロの中央矩形の奥行位置も増大したのに対して、反対コントラスト条件では奥行位置は出現せず一定のままであった。この結果は、T接合は面の統合の手がかりにならず面の統合には視差手がかりのみで十分なことが示された。
オクルード図形、オクルードされた図形および背景との関係は図10に図解されている。図の(a)には黒色縦帯のオクルード刺激の高さが灰色横帯の幅と同じにしたために、T接合が灰色横帯矩形に所属するかあるいは黒色縦帯矩形にあるのかを伝えないパターン、図(b)には黒色の縦帯を除いたために、T接合が無くなり代わりに横帯の断面の角と灰色背景との間に背景との境界を示すL接合が生ずるパターン、さらに図の(c)には横帯矩形の左右のエッジに対して背景を非交差視差にして奥行を出現させ、そのために横帯矩形に境界が無くなり、結果として3つの矩形面がひとつに視えるアモーダルな面の統合が成立するパターン、をそれぞれ示す。これを検証するために、図11に示したように、T接合の手がかりを弱めたステレオグラムで視差手がかりのみで面の統合知覚が生じるかが確かめられた。図の(a)は、同一コントラストでT接合を弱めたステレオグラムで、その図の中段の横帯の中央矩形は視差ゼロ、左端と中央あるいは中央と右端には交差あるいは非交差の勾配をもつ視差をつけた。このステレオグラムを両眼立体視すると下図のように中央の横帯が向こう側に傾斜して視える。図(b)は、コントラストを中段の横帯の中央と左右端矩形で反対にしたステレオグラムで両眼立体視すると下図のように中央の白色矩形は前額平行な独立した面として、同時にその左右には黒色傾斜面が視える。そこで、実験では視差要因による異なる面の統合におけるオクルーダーの幅の大きさの影響をしらべるために、オクル-ダーである縦帯の幅(0.1°、0.2°、0.4°、0.6°)および視差要因に関わる横帯の左・右端の幅(0.4°0.6°0.8°0.9°)が変えられた。横帯の左右端矩形視差を0、2.9、5.7、8.6、11.5′の5段階に変化し中央矩形の奥行位置を別に提示した奥行プローブ刺激とマッチングさせて測定した結果、視差の増大に連れて中央矩形の出現奥行は大きくなることを示したが、その奥行はオクルーダーである縦帯刺激の幅が0.2°を越えて広くなると小さくなること、また左・右端の幅のみを変化した場合にはそのような変化が生じなく視差に連動して奥行が変化することが示された。これはオクルーダー刺激の幅が大きくなると、面の統合効果が縮減されたことを示し、非オクルード面とオクルードの幅の比率(サポート比)が統合に関係していることを示唆した。そこで、サポート比と回帰直線の勾配(両端矩形の視差に対する中央矩形の奥行出現)をとると、その比率が大きいほど回帰勾配の直線が1.0に近似した。これは3次元の面の統合にはオクルーダーの幅の大きさのみが強く関与することを示した。
さらに、オクルードとオクルードされる刺激のパターン間の境界にズレを導入し、アモーダルな面の統合に及ぼす効果をしらべるために、図12に示したようなステレオグラムで実験した。ここでは、オクルード刺激である2本の縦帯の間の中央の矩形と左・右端との間に垂直方向のズレを導入し、左右端の矩形には視差を付けると共に同方向に運動する条件と静止条件を設けて、中央矩形の奥行位置をマッチング法で測定した。その結果、運動条件と静止条件ともズレの大きさが視角0.1度を超えると中央矩形の出現奥行が低下し、視角0.15度になるとほぼゼロとなった。3次元のアモーダルな面の統合はズレの幅が大きいと機能しなくなると考えられる。
また3次元の面の統合を、傾斜面を部分的にオクルードする事態を設定し、交差あるいは非交差視差条件でしらべた。実験で使用したステレオグラムは、図13にあるように、オクルード刺激を縦帯を1本にしたもので、黒色矩形と灰色縦帯の視差ゼロ(図a)および細い黒色縦帯刺激(視差勾配0.1°で45°傾斜面が出現)と灰色縦帯刺激(視差ゼロ)からなるステレオグラム(図b)を合体して構成したものである。このステレオグラムのコントラスト条件を、図cのように、横帯と縦帯が同一コントラストとなるように、あるいは図dのように、横帯刺激のうち、オクルーダーを挟んで左端と右端が反対のコントラストをとるようにそれぞれ設定した。同一コントラストの場合には図cの下図に示したように、面の統合作用が働かないと左端の面は鋭角に屈折して出現し、面の統合が働けば面全体が45°斜面として出現すると予測される。反対コントラスト条件では、図dの下図のように、面の統合作用が働かないのでひとつの面に統合されず、左端の面は鋭角に屈折して出現すると予測される。実験では、被験者に左端および中央矩形の奥行出現位置を上方の灰色矩形の奥行位置を変化させてマッチングさせた結果、同一コントラスト条件ではほぼ45°の面全体の斜面が出現し、反対コントラスト条件ではほぼ斜面は出現しなかった。これらの結果から、アモーダルな面の統合は単一のオクルード刺激からでも生起すること、また面が斜め角度をもっていても生起することが明らかにされた。
これら一連の実験結果から、(1)2つの縦帯のオクルード刺激によって3つに分断された横帯面の左右端矩形に視差を中央矩形には視差ゼロの3次元面事態では、アモーダルな面の統合は、分割面が同一の輝度コントラスト条件で成立し、反対コントラスト条件では成立しない、(2)横帯刺激の左右端矩形とオクルーダー刺激の境界にできる単眼奥行手がかりのT接合はアモーダルな面の統合に関与せず視差手がかりのみで十分である、(3)アモーダルな面の統合はアモーダルな2つの面の分離距離が大きくなると減じる、(4)アモーダルな面の統合はアモーダルな2つの面の間のズレが大きくなると減じる、(5) アモーダルな面の統合はオクルード刺激が単一でも生起し、その場合、出現する面は斜の角度をもつ面になる。
He et al.は、これらの結果から、アモーダルな面の統合過程についての2段階モデル(図14)を提唱している。第1段階は、2つの面の間の境界を割り当てるプロセス、第2段階は該当する面を拡大延長、そして統合する過程である。第1段階では、単眼手がかり(T接合)と両眼視手がかり(視差)によってオクルード面とオクルードされた面の間の境界の所属が決定される。第2段階では境界をもたない面が左右端から相互に作用しながらオクルード面の背後に伸びて広がる。その結果、3次元のアモーダルな面の統合が起きるというわけである。
錯覚的輪郭と輝度輪郭、および両要因の最適な組合せによるステレオグラム
Hartle et al.(11)は、Kanizsaタイプの主観的輪郭図形における輪郭面図形が奥行的に浮いて知覚される要因を探った。図15にあるように、主観的図形の輪郭を構成する面とその背後の地との間の輝度差を高い場合と低い場合とを設定し、誘導図形に視差だけをつけた条件と輝度差要因を組み合わせた条件で主観的面の奥行出現の程度に差が生じるかどうかを測定した。視差条件のみ、面と地の間の高低2つの輝度差条件、および視差と輝度差の組合条件の4通りのステレオグラムをミラー型のステレオスコープに提示し、すべての条件の誘導図形に0.09°の固定視差によって出現する凸曲面の頂点の高さを、別に提示したプローブの視差を変化(0.06°から0.17°)させるマッチング法で主観的等価値を測定した。
その結果、すべての条件での誘導図形の視差は固定してあるにも関わらず輝度差がある条件では、それが無い条件に比較して凸曲面頂点は有意に高く出現すること、また輝度差の高いあるいは低い条件間には有意差がないこと、さらに視差と輝度差の組合せ条件の凸曲面の出現の高さは視差のみ条件と低い輝度差条件の間になることが示された。これは、主観的輪郭を誘導する4つのエッジが輝度差による輪郭の凸曲面でオクルードされ、結果として凸曲面の出現を妨げるためと考えられる。輝度差による輪郭は、視差をつけた主観的輪郭の出現面をより高く知覚させる。
この結果をさらに確証するために、2者強制選択法で標準と比較刺激を提示し、凸曲面の高さの弁別の測定を4つの実験条件(視差のみ、視差と輝度差、低い輝度差、高い輝度差)内で実施した。標準刺激のステレオグラムの誘導図形視差は0.09°に固定し、比較刺激のそれは0.02°ステップで9段階に変化させた。その結果、低い輝度差の実験条件は他の条件に比較して閾値が有意に大きかった。もっとも閾値が小さかったのは、高い輝度差条件で、次に視差のみ条件と視差と輝度差の組合せ条件が続きほぼ同等の閾値を示した。
これらの結果は、ステレオグラムにおける主権的輪郭と輝度差による輪郭の組合せ条件における凸曲面の出現には、主観的輪郭がもつ本来的な多義性が影響することを示した。最尤推定法をこれらのデータに適用し、凸曲面知覚に及ぼす主観的輪郭と輝度差による輪郭の相対的効果を推定したところ、奥行知覚の標準偏差値は最適な手がかり組合せモデル(optimal cue combination model)に一致したが、その主観的等価値は主観的輪郭による効果を下回った。これは、手がかりの組合せに関して輝度による輪郭の規定する力より主観的輪郭の多義性による効果の方が大きいことを示唆する。
ステレオ視における面の連続と不連続
両眼視差は3次元世界における対象や対象の面の位置を高感度に検出できる。これが可能なのは、視覚システムが、網膜座標の両眼間の視差マップの対応関係を検出3次元視として知覚するからである(DeAngelis et al 1991)。しかし、最近の研究は両眼視差が徐々に連続して変化する場合、その周囲の視差情報に依存し奥行が減少して知覚されることを報告している(Cammack & Harris,2016; Deas & Wilcox, 2014, 2015)。Vreven et al. (2002)は2つの面の間にギャップを挿入したRDSの湾曲面で検出される視差の閾値をしらべると減少することも示した。Deas & Wilcox(2014)は視差検出による3次元面の視えの奥行視の減少には類似性や連続性などのゲシタルト要因が関係することを指摘した。3次元の奥行視が減じて知覚される現象をCammack & Harris(2016)は、対象を構成する視差を平均した値の差によって規定されているとした。たとえば、上方から凸面の台地を見た場合、それを構成する輪郭が台地の頂上まで地面からなだらかに構成されていると、その視差の平均値は地上か頂上まで垂直に立ち上がっている場合に較べて横幅は同等なので小さくなる。この視差の平均という視覚システムの機能が奥行視の程度を減じさせるというわけである。視差による3次元面の視えの奥行視のもう一つの説明は対象の面の連続と不連続、すなわち視差変化の急峻と面の断絶の相互作用によると考える。これまでの研究によれば、面の不連続のエッジは奥行知覚を促進し傾斜知覚閾値を改善するという結果(Cammack & Harris, 2016; Deas & Wilcox, 2014,2015, Wardle & Gillam, 2016)、あるいは逆にCraik-O’Brien-Cornsweet錯視の奥行の変形の場合には視えの奥行視の減少をもたらすという実験結果もある(Anstis, Howard, & Rogers, 1978;Rogers & Graham, 1983)。
そこで、Goutcher et al.(10)は、面の急峻(steepness)と不連続が組み合わされたときの奥行知覚への効果をしらべた。図16に示したように、ステレオグラムでは視差を操作して面の急峻(steepness)と連続・不連続が6通りに操作された。ここでは、標準刺激と比較刺激を用意し、実験1と2の比較刺激では面の急峻度を変化させ面の広さを一定(実験1)あるいは変化(実験2)させ、実験3と4の比較刺激では面の急峻度を変化させ中央部分にギャップを挿入(実験3)あるいは短い水平線分を挿入(実験4)させ、実験5と6の比較刺激では面の中央部分に奥行の不連続を挿入(実験5)あるいは中央部分に前額平行の不連続を挿入(実験6)した。ステレオグラムはランダムな白色ドットで作成し、各ドット間の視差は面全体の奥行距離と急峻度要因に依存して決められた。ステレオグラムは注視点と立体視を促す上下の縦線分(ノニウス線分)の後で2秒提示され、被験者(6名)には標準と比較刺激のステレオグラムの左右端のエッジ間の奥行距離を比較し、大きい方を選択させた。図17には実験1から6まで使用したステレオグラム(a、d、g)あるいはアナグリフステレオグラム(b、e、h)、そして面の急峻度(c、f、i)を示した。
実験の結果、(1)面のエッジからエッジまでの視差量が同一の場合、視差変化によって面の急峻度を増大させると視かけの奥行は小さい、(2) 面のエッジからエッジまでの視差量が同一の場合、面の急峻度が弱まる(視差11′、55′、110′)とエッジからエッジまでの奥行の平均主観的等価値(6.5、10.8、25.4arcmi)は増大する、(3)面の中央部分にギャップがあっても視差変化が急峻の場合には、ギャップがない条件と同様に視えの奥行が小さい(エッジからエッジまでの奥行の平均主観的等価値は視差11′、110′の時に6.6、31.1 arcmin)、(4) 面の中央部分をギャップの代わりに前額平行面を挿入しても視差変化が急峻の場合には同様に視えの奥行が小さい(奥行の平均主観的等価値は視差11′、110′の時に9.1、18 arcmi)、(5)面の エッジからエッジまでの間に2つの等価な奥行面のある条件では、視差が大きい場合に連続面条件より視えの奥行知覚は小さいこと(不連続が±2.2、±4.4 arcminのとき平均主観的等価値は16.4、18.9 arcmin)、(6)不連続面を横にずらした場合には視差を大きくしても視えの奥行は一定を保つ(横の不連続が0、±17.6、±35.2 arcminのとき平均主観的等価値は10.70、8.26、11.47 arcmin)ことなどが示された。
これらの結果を通覧すると、面のエッジからエッジまでの視差量が同一の場合、視えの奥行は視差の変化がゆるやかな場合よりも小さくなる。これは面に不連続があっても成り立つので、良い連続というゲシタルト原則はここでの視えの奥行視に関与していない。また、面の急峻の程度や不連続性おける奥行視は相対視差をグルーピングあるいは平均化するメカニズムに依存し、ハイパーキクロピアン(特定の視差変調の周波数チャンネルに選択的に反応)のフィルターリングによるスムーシング機能が連続する面の奥行知覚には影響しないと考えられる。すなわち、面のエッジからエッジまでの視えの奥行は相対視差の勾配が高い場合に大きく、この相対視差の勾配は面の不連続がある場合にとくに重要であり、ハイパーキクロピアン過程でのスムーシング効果では説明できないことを示唆する。 まとめると、ステレオ視パターンにおける視えの奥行の主観的等価値は相対的視差の統合によって規定され、視差が同一方向に急峻に変化する場合にはそのバイアスが大きいこと、逆に全体的な視差変化に抗する急峻な視差が挿入される場合にはそれによって視えの奥行は減じると考えられる。
自然シーンにおけるさまざまな奥行の変化とステレオ処理の課題
自然シーンにおける特定部分の奥行変化は固有の特徴をもつが、そのステレオ視に対する知覚的効果はいまだ明らかにされていない。自然シーンにおける奥行の変化は対象と均一では無い面の部分の間のエッジによっている。Iyer & Burge(14)は、自然シーンの奥行変化が両眼立体視のしくみ、とくに部分がオクルージョン(half occlusion)された事態の検出およびその視差検出にどのように影響するかを検討した。ここでは、自然シーンのステレオイメージおよびレーザーによる距離測量値が共に記載されたデータベースを用い、計算論による研究(computational study)が行われた。
まず、ステレオイメージからステレオイメージのパッチ(patch)をサンプリングし、それらのパッチから局所部分の奥行変化を示す視差コントラスト(中心視した位置の視差とそのごく近辺の視差disparity contrast)を量的に特定した。視差コントラストとは視差がゼロである中心視の位置のごく近辺の位置の視差を指す。その方法(図18)は、ステレオ3Dの幾何学に依拠し左右ステレオグラムイメージの対応点および視差値の補間手続である。図18の(A)では、左右眼の視線交差に生じる3Dシーンのサンプル点(白色の□)は左眼(LE)と右眼(RE)(眼球間距離65mm)に投影された輝度とレンジ(奥行距離)のイメージから得られることを示す。投影面(projection plane)上のサンプル点(pixel centers)は3Dシーンのポイントの部分集合となる。シーン内の3D surface(黒色の太線で表示)のサンプル点(白色の○)は左右眼の視線の交差するところとなる。視線上の小矢印は3D surface上の点からの反射光線を示し、各眼の輝度とレンジイメージでのpixel値(pixel value)を決める。3Dシーンのサーフェス上のサンプル点が3Dシーンのサンプル点と一致する場合もある(点線の円で囲んで表示)。この場合、これらの点からの光線は投影面のpixel centersで交差する。
図18の(B)には、左右眼イメージでの対応点の位置を決める手続きを示す。まずアンカーとした眼のイメージ(ここでは左眼)におけるピクセルセンターの位置をサンプル点(図中の1)として決める。これと対応する点(2)を左眼の3D surface上に決める。右眼上でこれと対応する点(3)を3D surfaceのサンプル点からレイトレーシングによって決める。右眼のイメージ上のもっとも近いpixel center(4)を選択する。このpixel centerと対応する右眼の3D surface上の点(5)を決める。左右眼それぞれの3D surfaceの点(2と5)から3Dシーンの点(6)を見つける。このサンプリングされた3Dシーンは、3D surface上の各点を通る左右眼の視線の交差するところとなる。左右眼の3D surfaceの各点とキクロピアンの眼の視線との交差する点(7)は左右眼の3D surface上の2つの点間のリニアな補間によって求められる。この補間よって求められた点から左右眼への光線(点線)が投影面と交差する点は左右眼の対応点(8)となる。この2つの対応点と3D surface上の補間した点との間の輻輳角(θ)が注視点となる。
図18の(C)は、補間の前のサンプリングエラーの起きる確率で、グラフ上の垂直点線は3Dシーンのサンプル点の間が一様に分布されていると仮定した場合を示す。図18の(D)は、補間後のサンプリングエラーの起きる確率である。
ハーフオクルージョンがある場合にはステレオグラムにおける対応点の算出にはエラーが起きる。そこで、ハーフオクルージョンにおける対応点算出の補間法を示す。図1 9は、ハーフオクルードの3Dシーンの各点、両眼視で観察できる3Dシーン上の各点および輻輳角が示されている。図19 の(A)には、 ハーフオクルードされた3D surface上の点を示し、この点(far 3D surface、黒色●で表示)は左眼では視えるが右眼ではオクルードされる。図中の矢印は補間に必要なレイトレーシングを示す。projection plane上の黒色■印は補間によって得られたイメージポイントを示す。3D surface上の点がハーフオクルードされた場合、これまでの補間法では無効な値となるので、 図19の(B)に示したように、両眼で観察できる3D surface上の点(黒色●で表示)とこれに対応するprojection plane上の2点(黒色■印)から輻輳角を推定する。surface上の点が両眼で観察できるとき求める輻輳角はどちらの側の眼がアンカーとなってもθとなる。その結果、図19 の(C)に示したように、求める輻輳角は3D surface上の点と左右眼のノーダルポイントで規定されたエピポーラー面内で計算される。このように視差による奥行変化を計算すると、自然シーンにおける視差の不連続(half occlusion)領域における対応点を求めることができる。
このような方法でステレオグラムにおける対応点を補間して求めた事例を図20に示した。図の(A)と(B)ともにステレオイメージに重ねて表示した左右ステレオペアの対応点(上段)および奥行距離のグラウンドトルース(groudtruth)データに重ねた対応点(下段)である。図の右(RE)と左(LE)ステレオグラムを平行視差観察、左(LE)と右(RE)ステレオグラムを交差観察する。左右イメージの3D surface上の各250の正しい対応点は黄色ドットで表示、ハーフオクルードされた対応点の候補あるいは無効の対応点は赤色ドットで表示してある。対応点を決定した後に輝度とレンジの値が対応点を中心とした一様なピクセルグリッド上で補間によって求められた。左右眼の輝度とレンジのステレオパッチはステレオイメージから得られる。グランドトルースの視差マップは左右のレンジイメージから直接計算される。
さらに注視点が曲面、コブ面あるいは傾斜面にある場合、その近傍の視差は変化する。この局所部分の視差変化(local depth variation)は、両眼で観察可能なシーンのステレオペアの中心点に関連づけて、以下のように計算される。
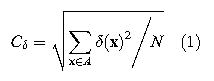
ここで、Cδ:local depth variation、δ:grundtruth relative disparity、A:local spatial integration area、X:空間位置、N:局所部分にあるピクセルの数
この式で計算されたステレオイメージが図21 (A)から(D)に視差コントラスト別に示されている。図の上段には自然シーンのステレオパッチ、下段にはこれに対応するグランドトルース視差マップである。局所部分の視差コントラスト(Cδ)は右側ステレオグラムになるにつれて大きくなる。各ピクセルでのグランドトルース視差(下段)はグランドトルースディスタンスから直接に計算し、色分けされた右端縦バーは視差(arcmin)を表し、センターピクセルにおける視差(輻輳角)に関連づけて表示されている。観察者がシーンのサーフェスのある1点を注視するかぎりステレオイメージパッチは左右で対応点をもつ。
Iyer & Burgeは、このように、ステレオイメージにおける対応点をサンプリングし、それらを補間することで視差を推定する方法を見いだした。その結果、自然シーンにおけるハーフオクルージョンとその視差の計算による検出が高度に信頼できること、また局所部分の視差変化(local depth variation) とハーフオクルージョンが有るか無いかの確率は網膜の離心率(eccentricity)にともなって変わること、さらに自然シーンにおけるハーフオクルージョンの分布の大きさと両眼視観察可能な領域の大きさの程度を確立した。 ここで示された自然シーンのあらゆる場合における視差検出は、ステレオビジョンの精神物理的研究の素材および自然シーンにおける視差の最適化のモデルになると考えられる。
両眼視におけるプリズム順応(シンメトリカルと非シンメトリカルプリズム条件の比較)
視差の融合バージェンスは速い(fast)成分および遅い(slow)成分から成立し、網膜視差を起動させる。またそれぞれの成分には神経生理的な時間差がある(Horwood & Riddell, 2008; McLin et al,1988; Schor, 1979; Semmlow & Wetzel, 1979)。速い成分の融合バージェンスとは、視対象の提示に際してバージェンス角度と視対象の間の視差を調整するための比較的速いそして正確な両眼間に共役性のない眼球運動が起きることをいう。遅い成分の融合バージェンスとは近いあるいは遠い視対象に時間をかけて順応する働きをいい、片眼が遮蔽されまた他の奥行手がかりがある場合にはこの働きで検出できる。この2つの成分は、異なる奥行位置にある対象を一つに融合するために相互作用する。さらに、この2つの融合バージェンス成分はプリズムをもちいることで変えられる。たとえば、眼球にベースアウトの向きでプリズムを装着すると眼球は内側に屈折すなわち輻輳(convergent)するし、逆にベースインの向きでプリズムを装着すると眼球を外側に屈折、すなわち拡散(divergent)させる。両眼視が1秒から15秒以上にわたると遅い成分の融合バージェンスが働きプリズムへの順応が起き、この人工的な斜視はプリズムを装着する前の正常なレベルに復帰する。この順応的な反応はプリズム順応(prism adaptation)と呼ばれる。プリズム順応の研究の多くは片側にプリズムを装着する条件(非シンメトリカル条件)でなされ、輻輳とサッカディク(sacadic)のシステムを刺激して装着側の網膜イメージがシフトされ、したがって視野も中心からシフトされる。両眼に同一のプリズムを装着(シンメトリカル条件)すると、正中線に沿って視野がシフトする。しかし、この種の2種類の異なる順応が健常な個人にも生じるかは不明である。
Santos et al.(31)は、シンメトリカルな斜視順応と非シンメトリカルなそれとが同一のしくみか否かを順応時間に応じて順応レベルがどのように変わるかをみることでしらべた。健常眼の被験者(16名、18から22歳齢)の両眼視能力について斜視の有無、輻輳の近点、プリズムに対する輻輳範囲などをしらべた上で、順応時間を変えて順応(0分から7分まで15段階)させ、その後直ちに順応効果をflashed Maddox Method(片眼に赤色ロッド、他眼に白色ロッドを提示し1眼に装着したプリズムの強度を変えてロッドがひとつに視える強度を測定)で水平方向の斜視の逸脱程度を測定した。被験者を座らせMuscle Imbalance Measureカードを40cm前方に提示し、垂直の赤色光線をフラッシュさせてその位置を報告させることでベースラインとなる斜視を初めに測定した。次に、被験者の両眼あるいは片眼にプリズムを装着し、順応時間後に斜視の逸脱程度を測定した。実験条件は4通り、非シンメトリカル条件で片眼に6Δのプリズムをベースインあるいはバースアウトで装着、およびシンメトリカル条件で両眼に3Δフリッパープリズムをベースインあるいはバースアウトでそれぞれ実験した図22)。
測定のデータから斜視順応比率(rate of phoria adaptation)を次式
(最後の斜視測定値-最初の斜視測定値)÷time constant(斜視順応が63%に安定する時間)
で算出し、各実験条件間で比較した。その結果、ベースアウトプリズム装着の非シンメトリカル条件の順応速度はシンメトリカル条件より有意に速かった。ベースインプリズム装着の非シンメトリカル条件の順応速度はシンメトリカル条件と有意差が無かった。これらのことは、遅い融合バージェンスの順応速度には方向性(輻輳あるいは開散)の非対称性があることを示した。多くの被験者では6Δのプリズムを7分順応後には完全に順応することが示されたが、これは4通りの実験条件間には相違がなかった。斜視レベルのもっとも完全な順応はベースアウトプリズム装着の非シンメトリカル条件で生じ、次はベースインプリズム装着の非シンメトリカル条件であった。また、輻輳あるいは開散が十分ではなく、または過度を示す被験者は健常な者と比較しシンメトリカル条件あるいは非シンメトリカル条件の斜視順応に対して顕著な相違が示された。これは臨床的な測定での融合バージェンスの範囲のデータとも関連すると示唆された。
近視者における時間的両眼情報の処理
両眼立体視のためには左右眼の入力像の空間的な属性、たとえば網膜像のピント度や空間周波数特性を知る必要があり、これらが両眼マッチング、視野闘争、両眼立体視力を規定するので多くの研究がある(Hoffman & Banks, 2010,Fahle, 1982, 1983; Shors, Wright, & Greene, 1992,Schor & Wood, 1983; Westheimer & McKee, 1980; Yang & Blake, 1991)。両眼視の空間的そして時間的特性の相互作用についてはいまだ知られていないが、両眼視の閾値は空間周波数が高いと影響されること、また視差自体の時間的変容によって変わることがわかっている (Lee, S., Shioiri, & Yaguchi,2003, 2007; Patterson, 1990,Kane et al.,2014; Nienborg et al., 2005; Norcia & Tyler, 1984)。しかし、同一の空間周波数で異なる2つのフリッカーイメージを用いた場合、両眼のバランスは時間的要因によってどのように変わるかは知られていない。
Vera-Diaz(39)は、両眼視のバランスが空間周波数を変えたとき時間的・空間周波数(TF)にどのように依存するかをしらべた。低空間周波数と高時間周波数は選択的に網膜神経節細胞の大細胞経路によって処理されるのに対して、高空間周波数と低時間周波数は選択的に網膜神経節細胞の小細胞経路によって処理(Nassi & Callaway 2009)されるので、この種の細胞の形態の相違は近視の問題においても感度の差をもたらすと考えられる。実験では、まずステレオプシスの閾値を測定し、次いで両眼バランステストを実施した。被験者は33名で平均年齢24.6であった。図23(A)には、シャッター眼鏡で立体視させステレオプシスの閾値を求めるスクリーンニングの手続きを示す。はじめに視差を減ずるように左から右に配置された小さなサークルのなかで前方に奥行出現してみえるもっとも視差の小さいサークルを選択させて閾値の大体の見当を被験者ごとにつけ、次に図(B)のように、2つの凹の奥行とひとつの凸の奥行からできている3つの円を被験者に示し、凸に視える奥行の円を選択させてステレオプシスの閾値が求められた。ピーク空間周波数は1、2、4、8c/deg、フリッカーの時間周波数は0、4、7.5、12 Hzとし、それぞれ左右眼で独立に変化させた。両眼視バランステストでは、被験者の左右眼に文字を提示するが、各眼のコントラストは右眼を100とし左眼は段階的に変え、被験者にはシャッター眼鏡をかけて提示された帯域通過(bandpass)をかけて表示した文字の各行を左から右へ読むように教示した。文字は1行に5文字を、4行にわたって提示する。1行の文字の空間周波数は固定されるが上の行から下の行になるにつれて空間周波数が増大された。1文字は左右各眼からの異なる1文字がオーバーラップして提示された(図24A)。図24(B)には、両眼間のバランス点は左眼に提示された文字の反応割合が0.5を示したときの両眼間コントラスト比(赤色の矢印)とし、被験者ごとにこの点を基準値して5段階にコントラスト比を変えた。被験者には、左から右に文字を読み上げるように教示した。
屈折異常を指標として14名の近視者(球面度数SEが平均-3.45±2.10DS)と19名の正視者(平均+0.27±0.29DS)に被験者を分けて実験の結果は考察され、ステレオプシスの閾値は正視者と比較して近視者で有意に大きいこと、とくに近視が強い者は閾値が高いこと、また時間周波数の増大に伴って閾値はリニアに大きくなるが近視者は常に閾値が大きくとくに4Hzで両グループの差が有意に大きいこといことが示された。さらに空間周波数の増大にともなって閾値はゆるやかに増大するが、ここでも近視者は正視者に較べて大きいことが示された。両眼視バランス事態での文字の読み取りテストでは、空間周波数あるいは時間周波数をそれぞれ独立に変化させたときの左右眼のコントラストバランスの値は近視者と正視者間で有意差がないこと、屈折の程度と両眼バランスの間にも有意な相関がないこと、しかし高空間周波数と低時間周波数では近視者は正視者より左右眼インバランスが高いことなどが示された。
これらの結果から、正視者と比較すると近視者はフリッカーした刺激に対してのステレオオプシスが良くなく、また高空間周波数と低時間周波数で左右眼インバランが大きいこと、しかし正視者の両眼視では空間周波数と時間周波数に同等に依存することが明らかにされた。
自然シーン観察における両眼視差の神経生理的反応
両眼視差の研究は、これまで刺激としてバーパターン、グレーティングパターンそしてRDSをコントロールし、そのメカニズムについて多くの成果を上げてきた((Backus et al.; 2001; Parker, 2007; Welchman, 2016)。一方、両眼立体視力(stereoacuity)の検査では、リアルな奥行を用いたテスト(Howard-Dolman apparatus、 Frisby Stereo Testなど)は人工的な刺激によるテストより閾値が小さいことが知られていた。これは、自然シーンの刺激パターンでは複数の奥行手がかりが存在するのに対し、人工的な刺激パターンでは両眼視差にのみ限定されているためである。したがって両眼視差のメカニズムを研究するためには、生態学的な意味をもつ自然シーンを刺激として用いる必要がある。 Duan et al.(7)は、128チャンネルの高密度脳波測定(high density EEG)を用い、2次元シーンにスクランブルをかけたイメージ、2次元自然シーンおよび3次元自然シーンを(図25)提示し、それらの事象関連電位を測定して比較し、両眼視差の固有な空間と時間の双方に関連した反応分布がどのようになるかをしらべた。刺激は、図26のように、スクランブルイメージを最初に、次に2次元自然シーン、そして3次元自然シーンというように連続して提示し、その間の事象関連電位を測定した。3次元視の提示は液晶シャッターを用い、2次元シーンおよびスクランブルイメージは両眼に同一の刺激が提示された。被験者には頭部に128個の電極を装着して提示される刺激パターンを観察させ、2番目と4番目の刺激が2次元あるいは3次元かをボタンで選択させた。EEGは0.1から50Hz の間で帯域濾過され、電極のインペーダンスは50kΩ以下に設定され、その位置はおもに前額と耳のまわりに装着された。
実験の結果、シーンの2次元あるいは3次元の識別は正答率が90%以上で、またその識別が正確な被験者は反応時間も速かいことが示された。この研究のメインのテーマである2次元自然シーンおよび3次元自然シーンにおけるそれぞれの視差関連電位を比較すると、マイナスの電位が持続的に観察され、視差固有の電位反応の大きさは観察者のステレオ視力と相関しているとともに、スクランブルシーンと2次元自然シーンとの反応の大きさとも異なっていた。3次元シーン内の対象の距離の大きさを白黒で表示したデプスマップ(depth map)を用いた視差事象関連電位も視差固有の大きな誘発電位が示された。さらに2次元シーンの事象関連電位は後頭頭頂葉でピークを示すのに対し、3次元シーンのそれは後方にシフトした。
反射性融合バージェンスの順応
バージェンス眼球運動は奥行をもつ注視点にシフトさせ、ステレオプシスのための眼球運動を調整する。バージェンス運動は眼球調節とカップリングし、バージェンス、眼球調節と注視点は3角連動の関係にある。最近の研究は反射性バージェンスの順応過程を明らかにし、その神経生理的しくみを探ろうとする。サッケードに類似したステップ状に連続するパルス(pulse-step) を与えると反射性バージェンスの運動をコントロールできる。この種のパルスは両眼の回転運動である輻輳(inward)あるいは開散(outward)を始動する。
Erkelens & Bobier (9)は、反射性の融合バージェンスの順応能力を順応過程の最中に反復提示したテスト刺激に対する眼球の運動の速度や振幅をダブルステップパラダイムで測定してしらべた。実験では白色の枠組のなかに白色の注視点を各眼に提示し、両眼間距離6cmおよび40cmの観察距離をとる場合の輻輳(8.44°の輻輳角)を視差に用い、シングルステップでベースラインフェーズを設定した。この視差位置からベースラインフェーズとは異なる視差刺激が間歇的に10回提示された(ダブルステップ)。図27Aに示すように、シングルステップ刺激は2°のステップで単独で視差提示し、ダブルステップ刺激は2°ステップ視差提示後175msあとに輻輳あるいは開散の視差をステップ±1.5°の範囲で変化して提示した。実験に際しては、図27Bのように、被験者毎にキャリブレーションを実施した後、「ベースラインフェーズ20回」-「順応フェーズ70回(double step、途中10回のテスト刺激を挿入)」-「回復フェーズ20回」の順序で刺激を左右眼にハプロスコープで提示した。「ベースラインフェーズ」では、輻輳あるいは開散条件で視差刺激を変えて(視差2°のステップ)を提示し、両眼視融合を確かめた。「順応フェーズ」では、視差2°からスタートし±1.5°の範囲で視差を変化して提示した。輻輳あるいは開散方向への視差の変化条件は、輻輳値増大(CGI)、輻輳値減少(CGD)、開散値増大(DGD)、開散値減少(DGI)の4通りとし、図のBにあるように提示した。輻輳あるいは開散に対する順応の程度をしらべるために視差2°ステップのテスト刺激を10回にわたって実験者制御によるオープンループで提示し、左右眼の単眼眼球運動を赤外線装置をを用いて記録した。
実験の結果、左右眼の輻輳の振幅と速度は輻輳値減少(CGD)と開散値減少(DGI)条件では順応フェーズにおいて輻輳値増大(CGI)条件より急速な変化が起きたが、開散値減少(DGI)条件では大部分の被験者の反応はたとえ変化があっても限定的だった。反射性の融合バージェンスの順応は輻輳と開散の増減で非対称の反応をすることが示された。
非交差視差に対する眼球の融合バージェンスの順応能力が限定されていることから、輻輳と開散の反射的融合バージェンスを担う神経過程は異なることを示唆する。これはVRを用いた場合など開散的融合(非交差示視差)で順応からの回復が遅いことの原因となる。